Launch your research career with a doctoral contract from Aix-Marseille University through ED62 or through NeuroSchool.

Launch your research career with a doctoral contract from Aix-Marseille University through ED62 or through NeuroSchool.
Launch your research career with a doctoral contract from Aix-Marseille University through ED62 or through NeuroSchool.

Translation starts with a Methionine: True, but not always, as revealed in a study of the PCP component Vangl2.
The brain: a permanent work in progress

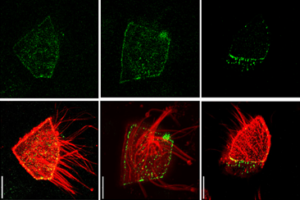
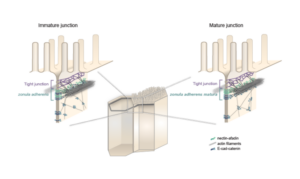
Lenne and Le Bivic teams show that intestinal adherens junctions are very different from the textbook picture.
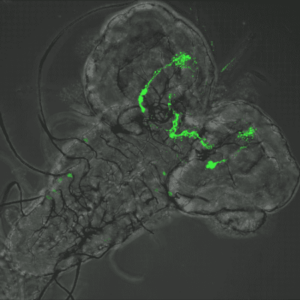
How the larval microbiota modulates the sensory modalities of future adults?
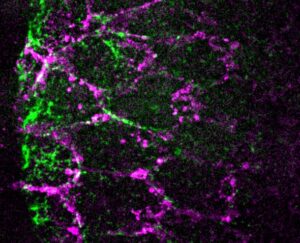
Understanding how adhesion and contractility drive a wave of tissue invagination from a subcellular perspective.
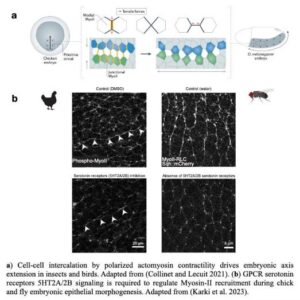
Sanjay Karki from Thomas Lecuit’s team reports that serotonin signaling regulates actomyosin contractility during embryonic axis morphogenesis in insect and bird.
Our team at the IBDM is part of a Maitre de Conference competition for a permanent assistant professor position in Bioinformatics in Aix-Marseille University.
Nous recherchons un/une gestionnaire administratif et financier (H/F). La personne recrutée sera intégrée au sein du Service Administratif et Financier de l’Institut de Biologie du Développement de Marseille (IBDM).
We are looking for candidates wishing to apply to a CENTURI Program PhD fellowship to work on the cell biology of epithelia in the emerging model Trichoplax.
Required: MSc in physics, biophysics, acoustics, molecular or cellular biology, neurosciences or a related discipline.

The work will investigate how transcription factor protein sequences influence LLPS, nuclear sub-compartmentalization and gene regulation during development.
The intern will develop skills in cell biology (cell culture, immunofluorescence, confocal imaging) and will be trained to use advanced biophysical tools (deformable hydrogel beads, atomic force microscopy).




