










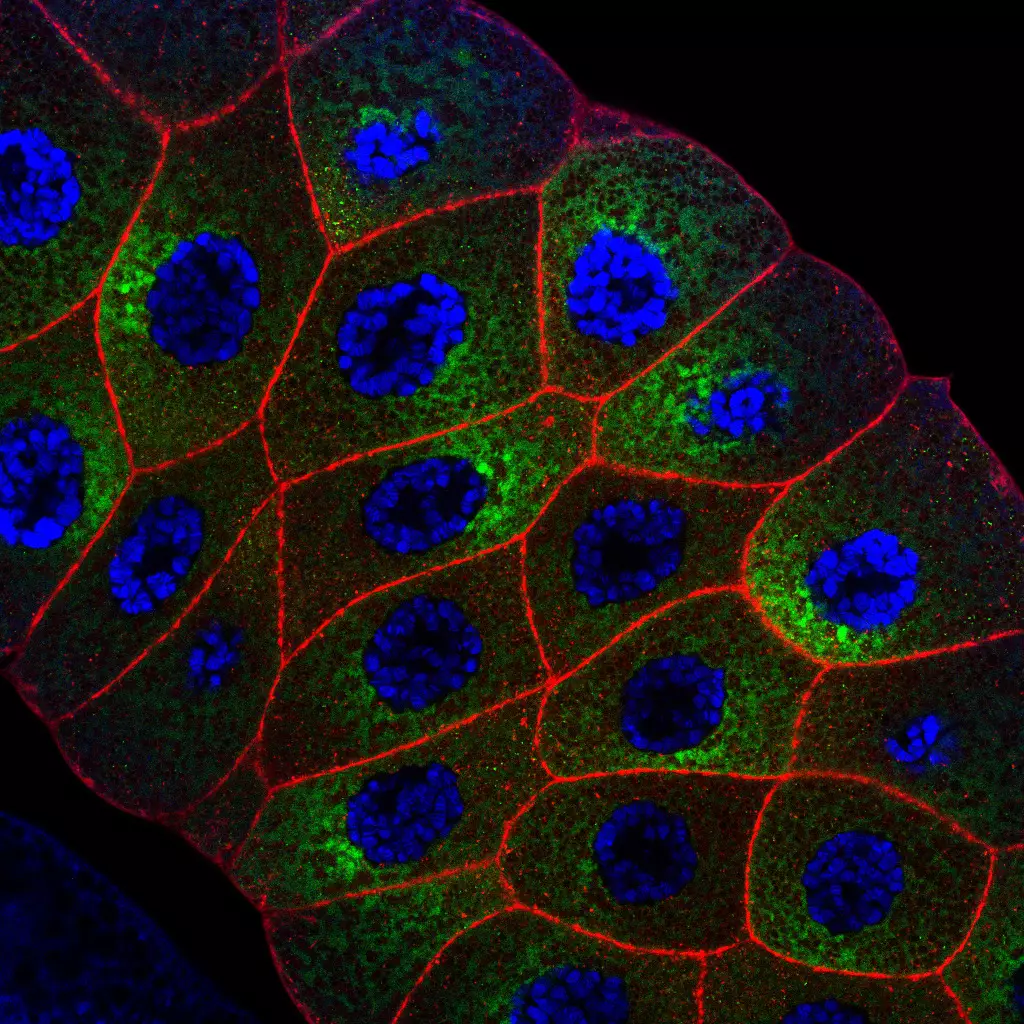
How the larval microbiota modulates the sensory modalities of future adults?
5 motivated and talented students successfully defended their thesis between September 2022 and January 2023.

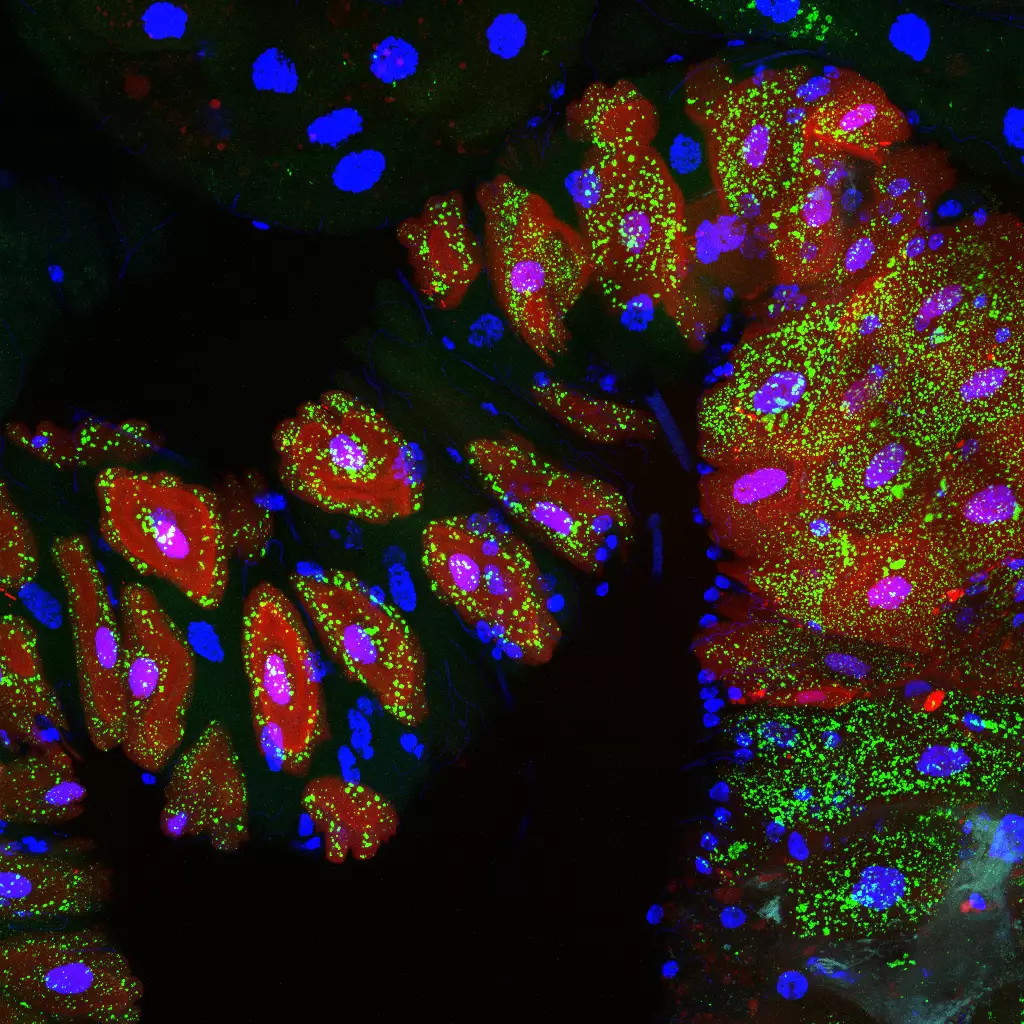
Our work demonstrates that by activating the NF-kB pathway, cell wall peptidoglycan produced by these same bacteria, counteracts SREBP activation in adipocytes in a cell-autonomous manner.
7 IBDM teams have received grants from ANR
7 IBDM teams have received grants from the Agence Nationale pour la Recherche (ANR) in 2021. Congratulations to Vincent Bertrand, Harold Cremer, Pascale Durbec, André Le Bivic, Pierre-François
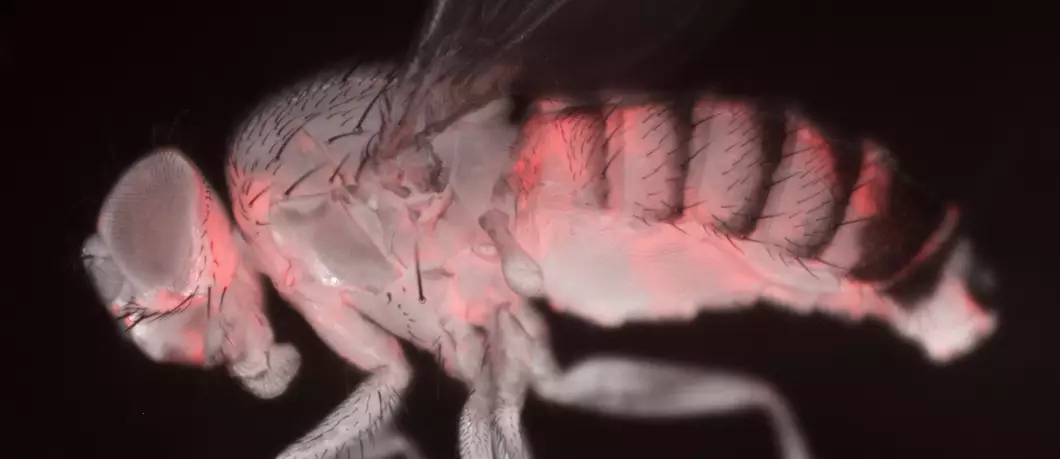
In a recent article published in iScience, Bernard Charroux, Fabrice Daian and Julien Royet analyze the behavior of fruit flies simply put in contact with bacteria potentially pathogenic to them.
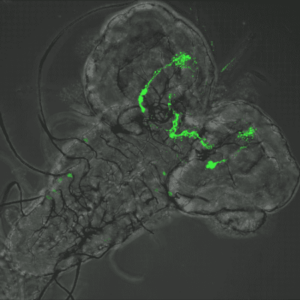
Julien Royet’s team uses the genetic tools generated in the fruit fly to dissect the molecular interactions between bacteria and the host’s nervous system.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}