
KL
Trainee
Kenza LARIBI
KL
YB
Researcher
MB
Trainee
MV
Msc Student
KK
Technical Staff
RC
Technical Staff
JG
PhD Student

Technical Staff


University Lecturer


University Lecturer


Researcher

Articles
Job opportunities
Articles
29/04/2022
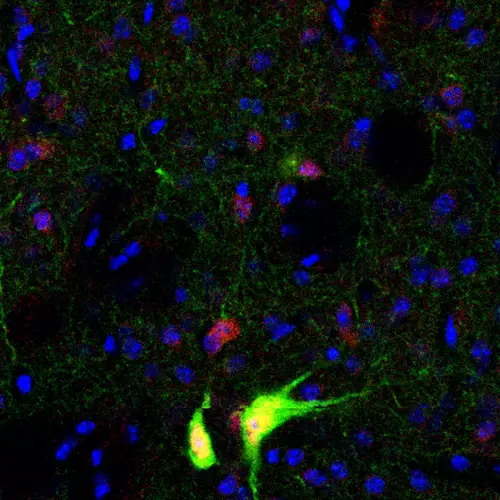
Targeted Tshz3 deletion in corticostriatal circuit components segregates core autistic behaviors
Nos résultats apportent un éclairage nouveau sur l’implication du circuit corticostrié dans les TSA.

Data published in Hum Mol Genet. confirm that Tshz3 heterozygous mice constitute a model that replicates many of the clinical problems reported in patients with TSHZ3 heterozygous conditions.
Job opportunities
Trainee
KL
Researcher
Trainee
Msc Student
Technical Staff
Technical Staff
PhD Student
Technical Staff
University Lecturer
University Lecturer
Researcher

Researcher

{kind=link}
{kind=link}
{kind=link}