
Hanyu Wang























Join the IBDM for your internship!
Seeking for your Master internship? The IBDM seems like the right place to do it? Check out our offers.

Congratulations to Robert Kelly, Frank Schnorrer, Cédric Maurange, Bianca Habermann and Delphine Delacour!

From AFM-Téléthon Postdoc Fellow to CNRS Researcher: Qiyan Mao is advancing human muscle research through molecular and tissue mechanics approaches.

IBDM Marseille inspires young minds: engaging primary school children on childhood cancer (“Contre le cancer, j’apporte ma pierre”) and interacting with high school students through immersive experiences (DECLICS).

Internal Seminar by Nuno Luis
Join us on 13/07/2023 at 12:30 in Amphi 12 for an exciting talk by Nuno Luis from our Team!
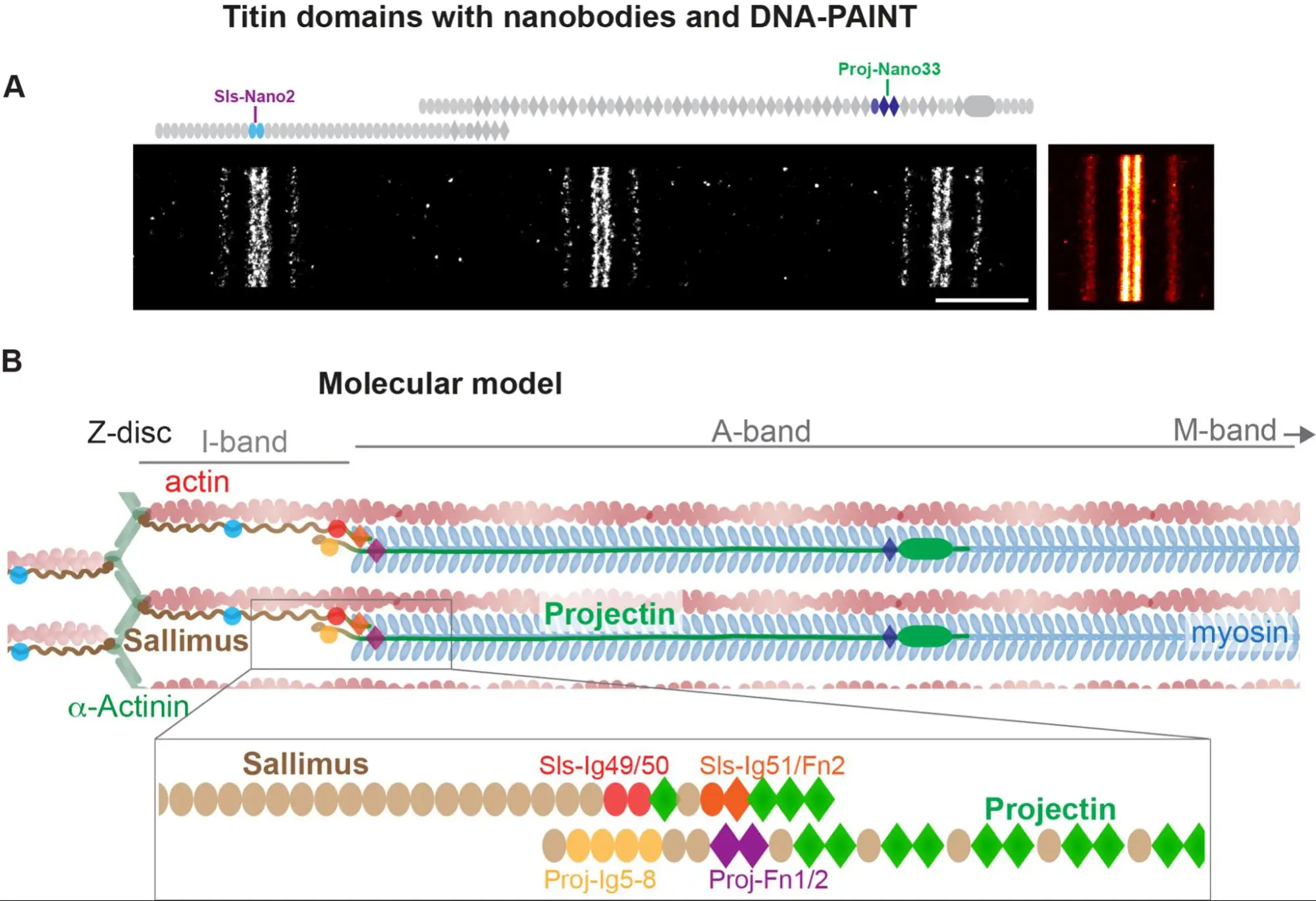
Drosophila sarcomeres visualised at super-resolution with the help of nanobodies and single molecule blinks.
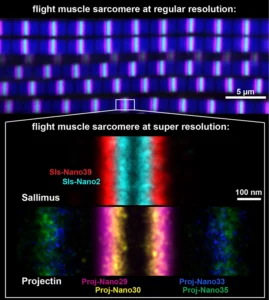
Organizing the organizers
A new perspective to tackle the very complex cell polarity field.
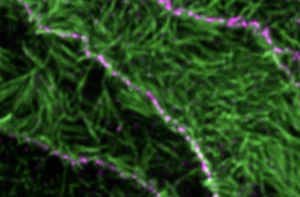
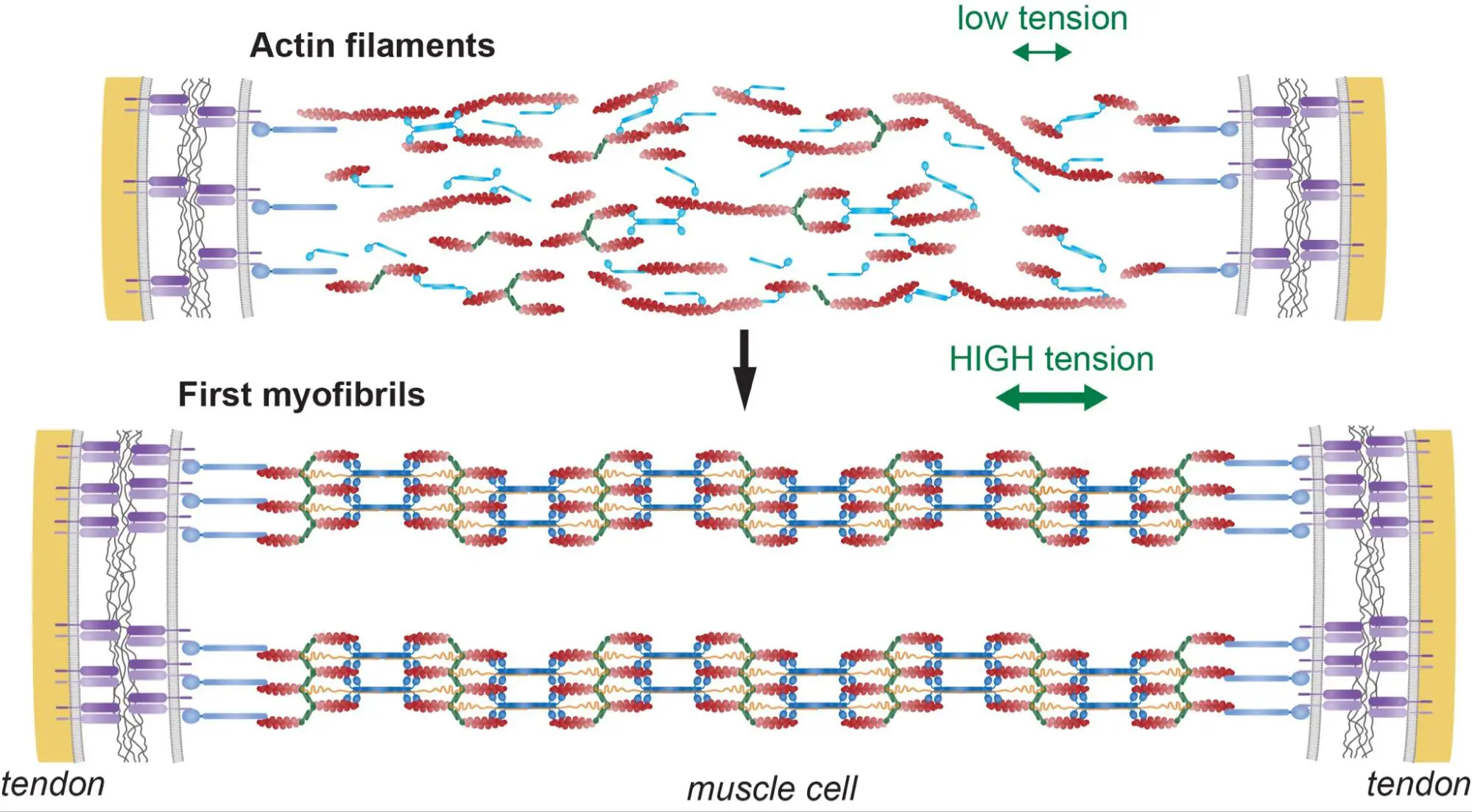
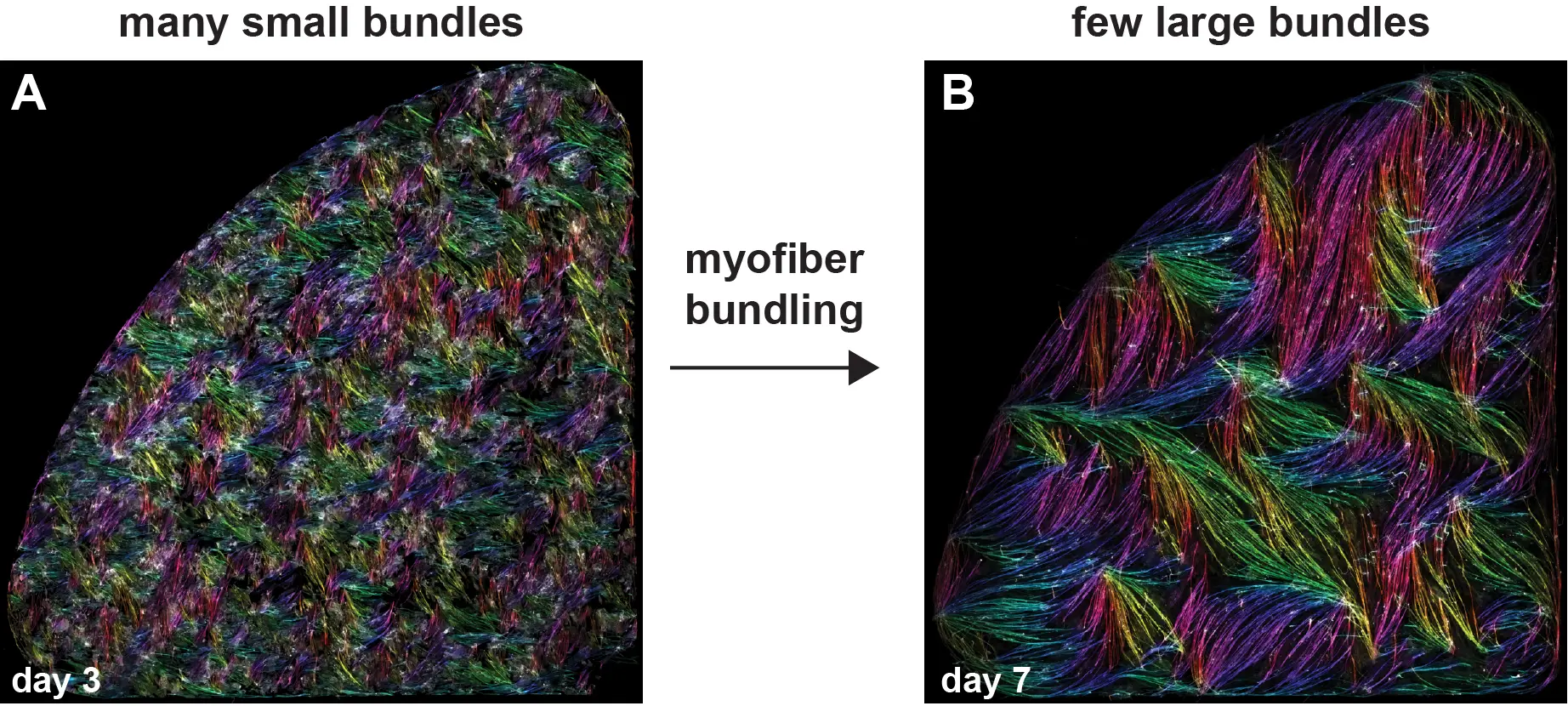
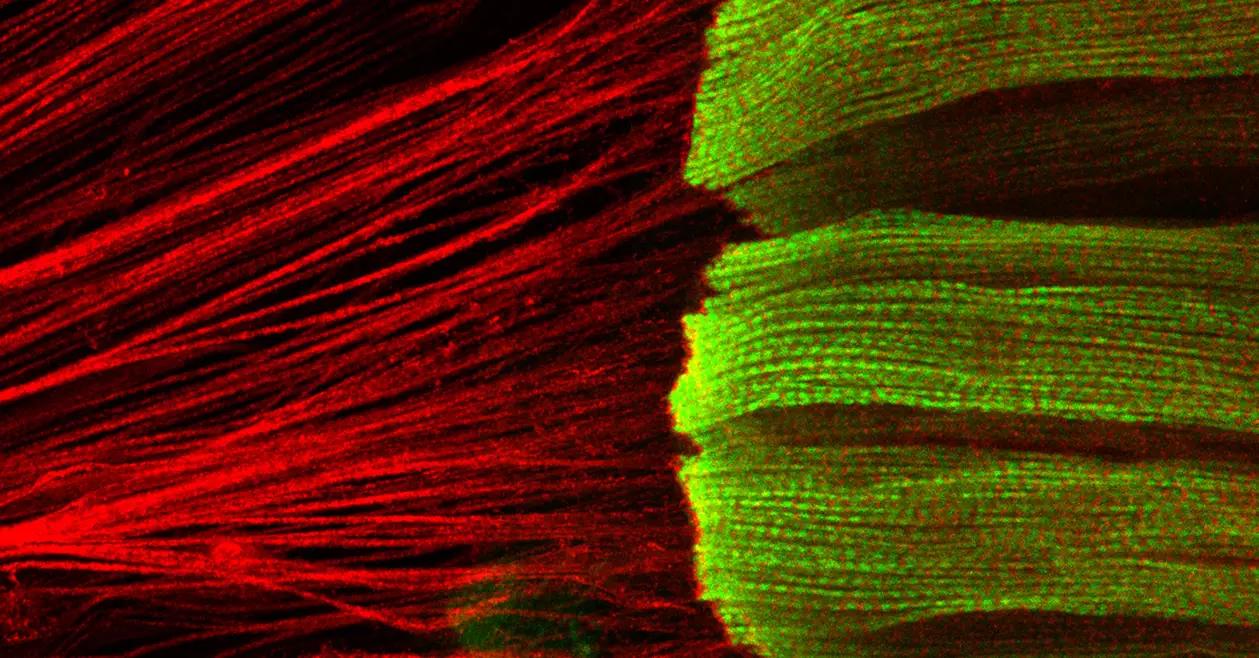
Self-organisation of human muscles in a dish
Human muscle cells self-organise into defined fiber bundles in vitro even without the presence of external cues !
Frank Schnorrer elected as EMBO member
EMBO elects 67 new members and associate members. They join the community of more than 1,900 leading life scientists in Europe and beyond.

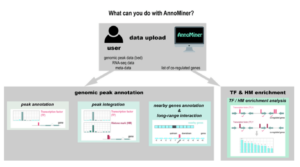
We introduce a novel, user-friendly web-based tool ‘AnnoMiner’ to annotate and integrate epigenetic and transcription factor binding data.
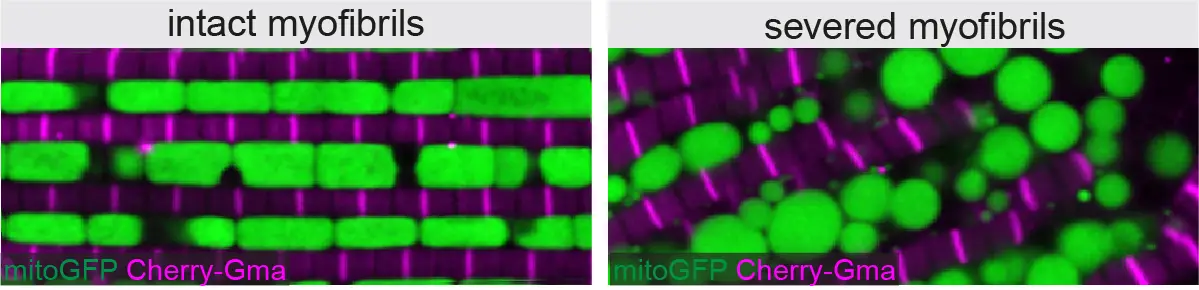
By combining genetics in the fruit fly Drosophila with state-of-the-art imaging and deep-learning, researchers at IBDM have found that mitochondria coordinate their formation with myofibril development to match the correct muscle type.
The Hippo pathway controls muscle growth
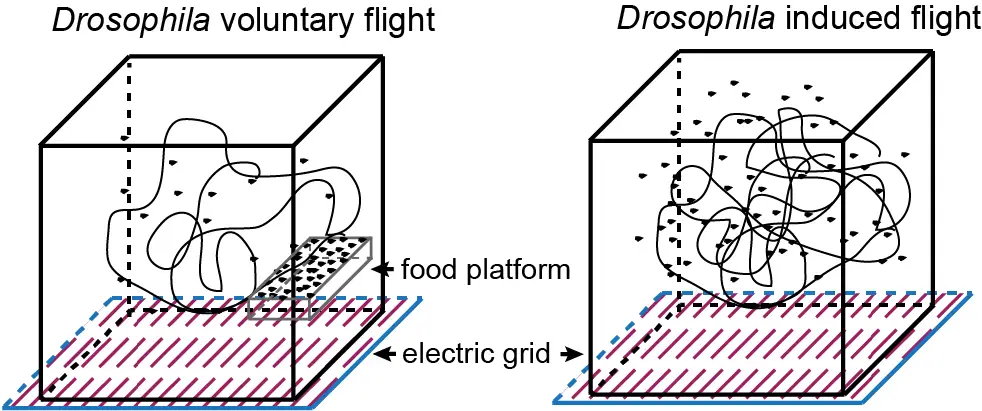
Schnorrer team and colleagues discovered that a signalling pathway, called the Hippo pathway, is controlling muscle growth during development of Drosophila flight muscles.
European Synergy Grant for Muscle Research
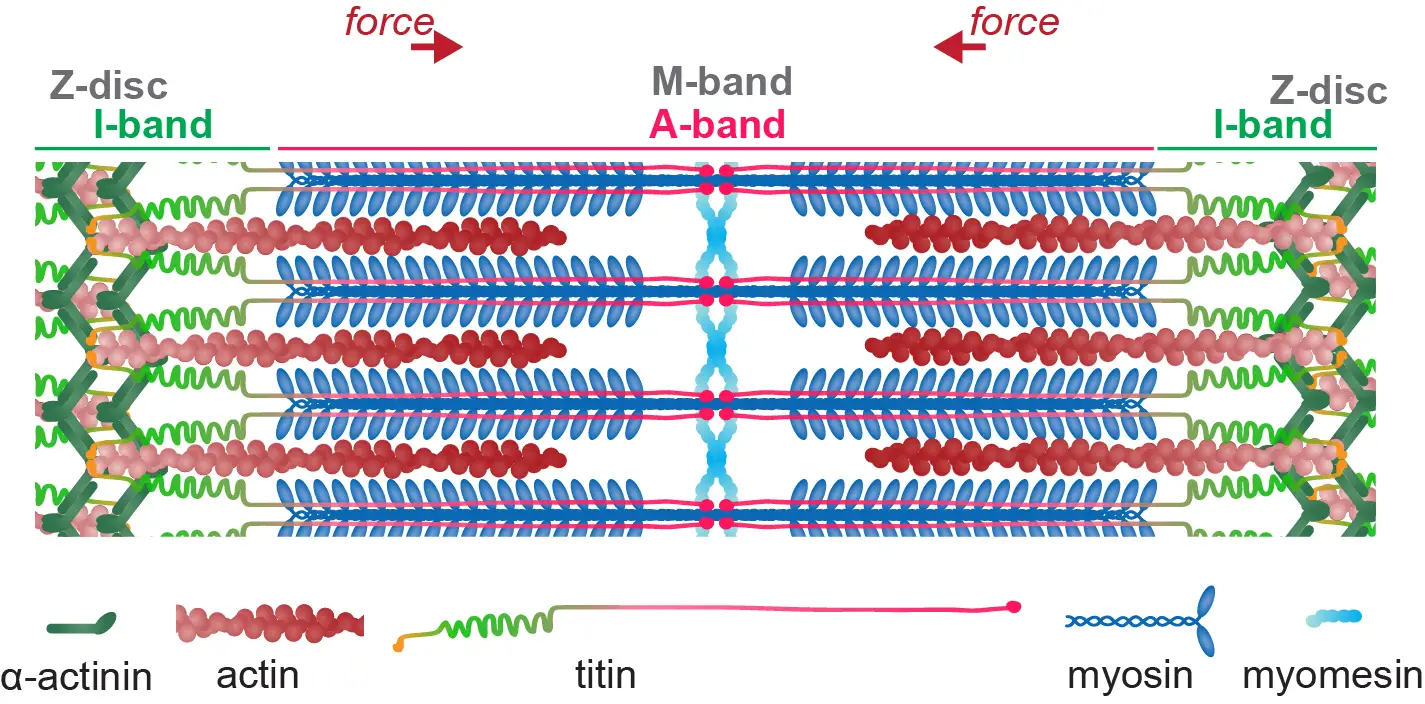
The European Research Council (ERC) awarded one of the rare ERC Synergy Grants to an international consortium of scientists, Frank Schnorrer, Stefan Raunser, Dirk Görlich and Mathias Gautel.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}