Lancez votre carrière de chercheur avec un contrat doctoral à l’Université Aix-Marseille par le biais de l’ED62 ou de la NeuroSchool.

Lancez votre carrière de chercheur avec un contrat doctoral à l’Université Aix-Marseille par le biais de l’ED62 ou de la NeuroSchool.
Lancez votre carrière de chercheur avec un contrat doctoral à l’Université Aix-Marseille par le biais de l’ED62 ou de la NeuroSchool.

Dans les embryons de C. elegans les ligands Wnt diffusent dans le tissu pour polariser des cellules à distance.
La traduction commence par une Méthionine: Vrai, mais pas toujours, comme le montre une étude sur Vangl2.
Le cerveau : un chantier permanent

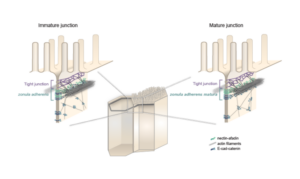
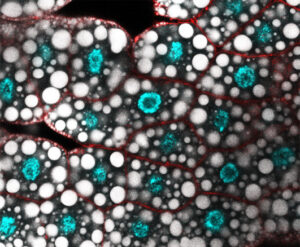
Les équipes Lenne et Le Bivic démontrent une nouvelle organisation pour les jonctions d’adhérence intestinales.
Comment le microbiote larvaire influence-t-il les modalités sensorielles des futurs adultes ?
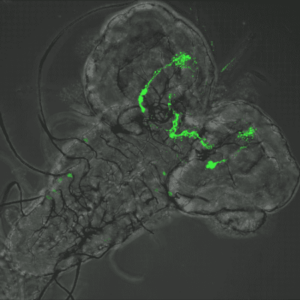
Comment l’adhésion et la contractilité entraînent une vague d’invagination tissulaire d’un point de vue subcellulaire.
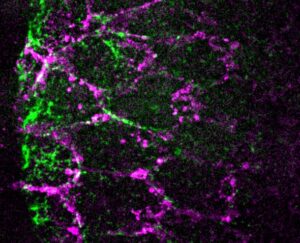
How two recently discovered and oppositely acting transcriptional regulators control metabolism in the Drosophila larval fat body, with a special attention on metabolic paths linked to fat accumulation.
Notre équipe à l’IBDM participe à un concours de maître de conférence pour un poste de professeur assistant permanent en bio-informatique à Aix-Marseille Université.
Nous recherchons un/une gestionnaire administratif et financier (H/F). La personne recrutée sera intégrée au sein du Service Administratif et Financier de l’Institut de Biologie du Développement de Marseille (IBDM).
We are looking for candidates wishing to apply to a CENTURI Program PhD fellowship to work on the cell biology of epithelia in the emerging model Trichoplax.
Conditions requises : Maîtrise en physique, biophysique, acoustique, biologie moléculaire ou cellulaire, neurosciences ou dans une discipline connexe.

Les travaux porteront sur la manière dont les séquences de protéines de facteurs de transcription influencent la séparation de phase liquide-liquide (LLPS), la sous-compartimentation nucléaire et la régulation des gènes au cours du développement.