









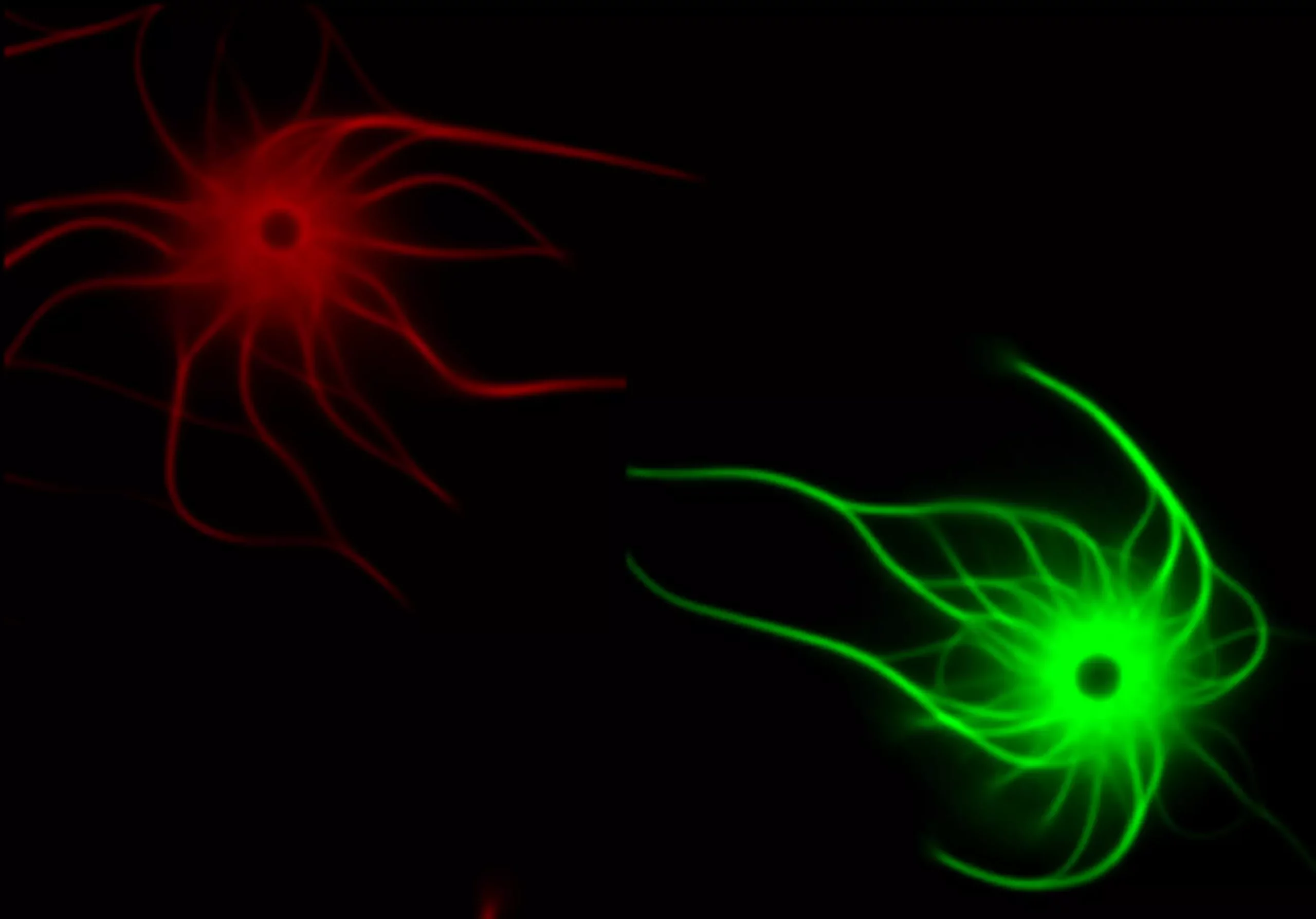
Un mécanisme pour expliquer la spécialisation d’isoformes d’actines malgré leur similarité
Nos résultats permettent de proposer un modèle simple et efficace sur la façon dont le vivant a pu générer de nouvelles fonctions grâce à l’utilisation de nouveaux isoformes d’actine.
L’équipe d’Alphée Michelot, en collaboration avec l’équipe de Pekka Lappalainen (Université d’Helsinki), a identifié et caractérisé une famille d’analogues nucléotidiques fluorescents très sensibles et structurellement compatibles avec l’actine.
Mechanical stiffness of reconstituted actin patches correlates tightly with endocytosis efficiency
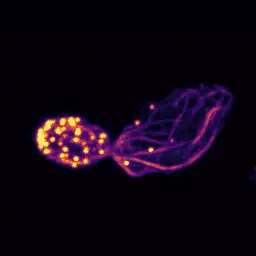
Une collaboration multi-disciplinaire entre l’équipe d’Alphée Michelot et l’équipe d’Olivia du Roure et de Julien Heuvingh à l’ESPCI publie dans PLoS Biology une étude montrant que la rigidité mécanique des patchs endocytiques d’actine est fortement liée à l’efficacité de l’endocytose.
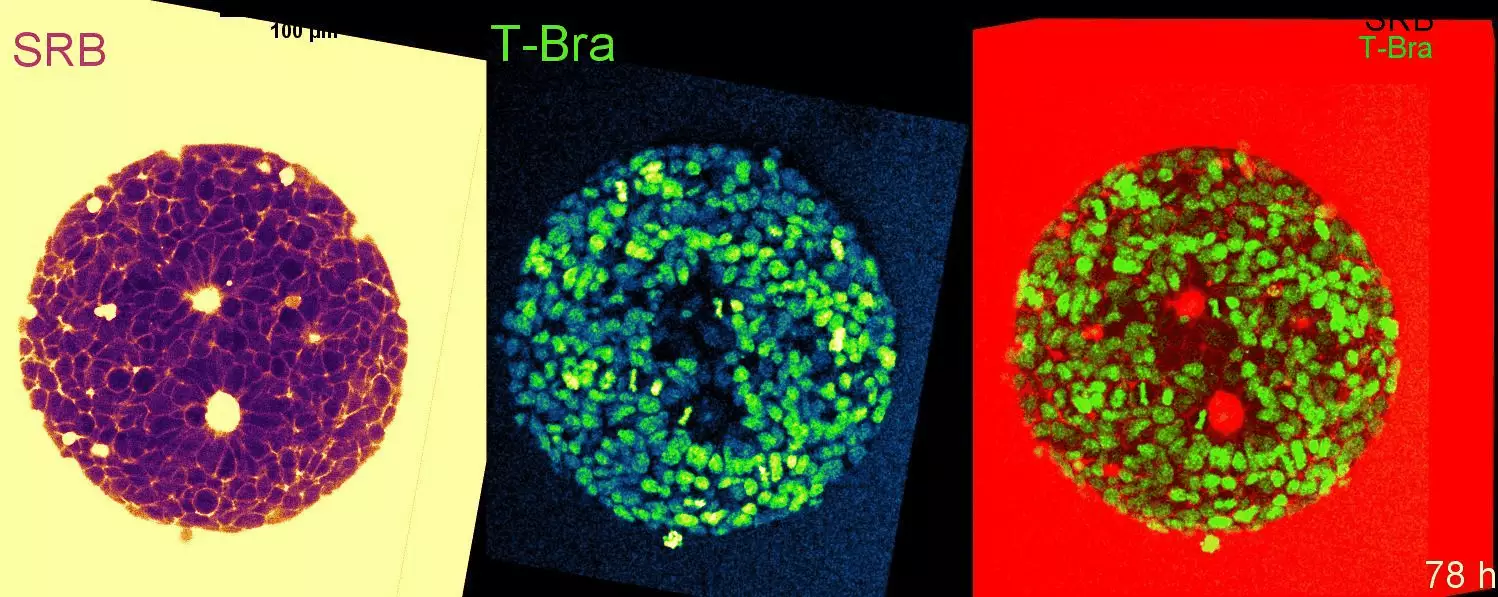
L’équipe d’Alphée Michelot publie dans Plos Biology un article expliquant comment les cellules contrôlent la taille de multiples réseaux d’actine au sein d’un cytoplasme commun.



{kind=link}
{kind=link}